In neuroanatomy, the optic chiasm, or optic chiasma ( /ɒptɪk kaɪæzəm/; from Greek χίασμα 'crossing', from Ancient Greek χιάζω 'to mark with an X'), is the part of the brain where the optic nerves cross. It is located at the bottom of the brain immediately inferior to the hypothalamus.[1] The optic chiasm is found in all vertebrates, although in cyclostomes (lampreys and hagfishes), it is located within the brain.[2][3]
| Optic chiasm | |
|---|---|
 Brain viewed from below; the front of the brain is above. Visual pathway with optic chiasm (X shape) is shown in red (image from Andreas Vesalius' Fabrica, 1543). | |
 Optic nerves, chiasm, and optic tracts | |
| Details | |
| System | Visual system |
| Function | Transmit visual information from the optic nerves to the occipital lobes of the brain |
| Identifiers | |
| Latin | chiasma opticum |
| MeSH | D009897 |
| NeuroNames | 459 |
| NeuroLex ID | birnlex_1416 |
| TA98 | A14.1.08.403 |
| TA2 | 5668 |
| FMA | 62045 |
| Anatomical terms of neuroanatomy | |
This article is about the optic chiasm of vertebrates, which is the best known nerve chiasm, but not every chiasm denotes a crossing of the body midline (e.g., in some invertebrates, see Chiasm (anatomy)). A midline crossing of nerves inside the brain is called a decussation (see Definition of types of crossings).
Structure
editIn all vertebrates, the optic nerves of the left and the right eye meet in the body midline, ventral to the brain. In many vertebrates the left optic nerve crosses over the right one without fusing with it.[4]
In vertebrates with a large overlap of the visual fields of the two eyes, i.e., most mammals and birds, but also amphibians, reptiles such as chameleons, the two optic nerves merge in the optic chiasm. In such a merged optic chiasm, part of the nerve fibres do not cross the midline, but continue towards the optic tract of the ipsilateral side. By this partial decussation, the part of the visual field that is covered by both eyes is fused so that the processing of binocular depth perception by stereopsis is enabled (see Figure 2).
In the case of such partial decussation, the optic nerve fibres on the medial sides of each retina (which correspond to the lateral side of each visual hemifield, because the image is inverted) cross over to the opposite side of the body midline. The inferonasal retina are related to the anterior portion of the optic chiasm whereas superonasal retinal fibers are related to the posterior portion of the optic chiasm.
The partial crossing over of optic nerve fibres at the optic chiasm allows the visual cortex to receive the same hemispheric visual field from both eyes. Superimposing and processing these monocular visual signals allow the visual cortex to generate binocular and stereoscopic vision. The net result is that the right cerebral hemisphere processes left visual hemifield, and the left cerebral hemisphere processes the right visual hemifield.
Beyond the optic chiasm, with crossed and uncrossed fibers, the optic nerves are called optic tracts. The optic tract inserts on the optic tectum (in mammals known as superior colliculus) of the midbrain. In mammals they also branch off to the lateral geniculate body of the thalamus, in turn giving them to the occipital cortex of the cerebrum.[5]
Arterial supply
editThe optic chiasma receives its arterial supply from the anterior cerebral arteries, and from branches of the internal carotid artery which ascend along the pituitary stalk (the latter supplying the midline portion of the chiasma).[6]
Development in mammals
editDuring development, the crossing of the optic nerves is guided primarily by cues such as netrin, slit, semaphorin and ephrin; and by morphogens such as sonic hedgehog (Shh) and Wnt.[7] This navigation is mediated by the neuronal growth cone, a structure that responds to the cues by ligand-receptor signalling systems that activate downstream pathways inducing changes in the cytoskeleton.[8] Retinal ganglion cell (RGC) axons leaving the eye through the optic nerve are blocked from exiting the developing pathway by Slit2 and Sema5A inhibition, expressed bordering the optic nerve pathway. Ssh expressed at the central nervous system midline inhibits crossing prior to the chiasm, where it is downregulated.[9][10] The organization of RGC axons changes from retinotopic to a flat sheet-like orientation as they approach the chiasm site.[11]
Most RGC axons cross the midline at the ventral diencephalon and continue to the contralateral superior colliculus. The number of axons that do not cross the midline and project ipsilaterally depends on the degree of binocular vision of the animal (3% in mice and 45% in humans do not cross).[9] Ephrin-B2 is expressed at the chiasm midline by radial glia and acts as a repulsive signal to axons originating from the ventrotemporal retina expressing EphB1 receptor protein, giving rise to the ipsilateral, or uncrossed, projection.[9] RGC axons that do cross at the optic chiasm are guided by the vascular endothelial growth factor, VEGF-A, expressed at the midline, which signals through the receptor Neuropilin-1 (NRP1) expressed on RGC axons.[12] Chiasm crossing is also promoted by Nr-CAM (Ng-CAM-related cell adhesion molecule) and Semaphorin6D (Sema6D) expressed at the midline, which form a complex that signals to Nr-CAM/Plexin-A1 receptors on crossing RGC axons.[13]
Other animals
editMammals
editSince all vertebrates, even the earliest fossils[14] and modern jawless ones,[5] possess an optic chiasm, it is not known how it evolved.[15] A number of theories have been proposed for the function of the optic chiasm in vertebrates (see theories). According to the Axial Twist theory the optic chiasm develops as a consequence of a twist in the early embryo.[16]
In Siamese cats with certain genotypes of the albino gene, the wiring is disrupted, with more of the nerve-crossing than normal.[17] Since siamese cats, like albino tigers, also tend to cross their eyes (strabismus), it has been proposed that this behavior might compensate the abnormal amount of decussation.[18][19]
Cephalopods and insects
editIn cephalopods and insects the optic tracts do not cross the body midline, so each side of the brain processes the ipsilateral eye.
History
editThe crossing of nerve fibres, and the impact on vision that this had, was probably first identified by Persian physician "Esmail Jorjani", who appears to be Zayn al-Din Gorgani (1042–1137).[20]
Additional images
edit-
 Scheme showing central connections of the optic nerves and optic tracts.
Scheme showing central connections of the optic nerves and optic tracts. -
 Brain seen from below, with the optic chiasm seen in yellow in the centre.
Brain seen from below, with the optic chiasm seen in yellow in the centre. -
 Transformations of the visual field toward the visual map on the primary visual cortex.
Transformations of the visual field toward the visual map on the primary visual cortex. -
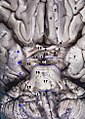 Brain and brainstem seen from below
Brain and brainstem seen from below -
 Left hemisphere of the brain seen in a cadaveric specimen from the side, with the optic chiasm labelled.
Left hemisphere of the brain seen in a cadaveric specimen from the side, with the optic chiasm labelled. -
 Cerebrum, inferior view, deep dissection.
Cerebrum, inferior view, deep dissection. -
 Guidance of axon crossing and non-crossing during development.
Guidance of axon crossing and non-crossing during development.







See also
editReferences
edit- ^ Colman, Andrew M. (2006). Oxford Dictionary of Psychology (2nd ed.). Oxford University Press. p. 530. ISBN 978-0-19-861035-9.
- ^ Bainbridge, David (30 June 2009). Beyond the Zonules of Zinn: A Fantastic Journey Through Your Brain. Harvard University Press. p. 162. ISBN 978-0-674-02042-9. Retrieved 22 November 2015.
- ^ de Lussanet, Marc H.E.; Osse, Jan W.M. (2012). "An ancestral axial twist explains the contralateral forebrain and the optic chiasm in vertebrates". Animal Biology. 62 (2): 193–216. arXiv:1003.1872. doi:10.1163/157075611X617102. ISSN 1570-7555. S2CID 7399128.
- ^ Stephen, Polyak (1957). The vertebrate visual system. Chicago: Chicago Univ. Press.
- ^ a b Nieuwenhuys, R.; Donkelaar, H.J.; Nicholson, C.; Smeets, W.J.A.J.; Wicht, H. (1998). The central nervous system of vertebrates. New York: Springer. ISBN 9783642621277.
- ^ Standring, Susan (2020). Gray's Anatomy: The Anatomical Basis of Clinical Practice (42th ed.). New York: Elsevier. p. 420. ISBN 978-0-7020-7707-4. OCLC 1201341621.
- ^ Erskine, L.; Herrera, E. (2007). "The retinal ganglion cell axon's journey: Insights into molecular mechanisms of axon guidance". Developmental Biology. 308 (1): 1–14. doi:10.1016/j.ydbio.2007.05.013. hdl:10261/338550. PMID 17560562.
- ^ Gordon-Weeks, PR (2005). Neuronal Growth Cones. Cambridge University Press. ISBN 9780511529719.
- ^ a b c Herrera, E; Erskine, L; Morenilla-Palao, C (2019). "Guidance of retinal axons in mammals". Seminars in Cell & Developmental Biology. 85: 48–59. doi:10.1016/j.semcdb.2017.11.027. PMID 29174916. S2CID 24381059.
- ^ Rasband, Kendall; Hardy, Melissa; Chien, Chi-Bin (2003). "Generating X, Formation of the Optic Chiasm". Neuron. 39 (6): 885–888. doi:10.1016/S0896-6273(03)00563-4. PMID 12971890.
- ^ Guillery, RW; Mason, CA; Taylor, JS (1995). "Developmental determinants at the mammalian optic chiasm". The Journal of Neuroscience. 15 (7): 4727–4737. doi:10.1523/JNEUROSCI.15-07-04727.1995. PMC 6577905. PMID 7623106.
- ^ Erskine, L; Reijntjes, S; Pratt, T (2011). "VEGF signaling through neuropilin 1 guides commissural axon crossing at the optic chiasm". Neuron. 70 (5): 951–965. doi:10.1016/j.neuron.2011.02.052. PMC 3114076. PMID 21658587.
- ^ Kuwajima, T; Yoshida, Y; Pratt, T (2012). "Optic chiasm presentation of Semaphorin6D in the context of Plexin-A1 and Nr-CAM promotes retinal axon midline crossing". Neuron. 74 (4): 676–690. doi:10.1016/j.neuron.2012.03.025. PMC 3361695. PMID 22632726.
- ^ Janvier, P. (1996). Early vertebrates. New York: Clarendon Press, Oxford University Press. ISBN 978-0198540472.
- ^ de Lussanet, M.H.E.; Osse, J.W.M. (2012). "An ancestral axial twist explains the contralateral forebain and the optic chiasm in vertebrates". Animal Biology. 62 (2): 193–216. arXiv:1003.1872. doi:10.1163/157075611X617102. S2CID 7399128.
- ^ de Lussanet, M.H.E. (2019). "Opposite asymmetries of face and trunk and of kissing and hugging, as predicted by the axial twist hypothesis". PeerJ. 7: e7096. doi:10.7717/peerj.7096. PMC 6557252. PMID 31211022.
- ^ Schmolesky MT, Wang Y, Creel DJ, Leventhal AG (2000). "Abnormal retinotopic organization of the dorsal lateral geniculate nucleus of the tyrosinase-negative albino cat". J Comp Neurol. 427 (2): 209–19. doi:10.1002/1096-9861(20001113)427:2<209::aid-cne4>3.0.co;2-3. PMID 11054689. S2CID 32536933.
- ^ Guillery, RW; Kaas, JH (June 1973). "Genetic abnormality of the visual pathways in a "white" tiger". Science. 180 (4092): 1287–9. Bibcode:1973Sci...180.1287G. doi:10.1126/science.180.4092.1287. PMID 4707916. S2CID 28568341.
- ^ Guillery RW (May 1974). "Visual pathways in albinos". Sci. Am. 230 (5): 44–54. Bibcode:1974SciAm.230e..44G. doi:10.1038/scientificamerican0574-44. PMID 4822986.
- ^ Davis, Matthew C.; Griessenauer, Christoph J.; Bosmia, Anand N.; Tubbs, R. Shane; Shoja, Mohammadali M. (2014-01-01). "The naming of the cranial nerves: A historical review". Clinical Anatomy. 27 (1): 14–19. doi:10.1002/ca.22345. ISSN 1098-2353. PMID 24323823. S2CID 15242391.
External links
edit- "Anatomy diagram: 13048.000-1". Roche Lexicon - illustrated navigator. Elsevier. Archived from the original on 2014-11-07.