Choristodera (from the Greek χωριστός chōristos + δέρη dérē, 'separated neck'[2]) is an extinct order of semiaquatic diapsid reptiles that ranged from the Middle Jurassic, or possibly Triassic, to the Miocene (168 to 20 or possibly 11.6 million years ago). Choristoderes are morphologically diverse, with the best known members being the crocodile-like neochoristoderes such as Champsosaurus. Other choristoderans had lizard-like or long necked morphologies. Choristoderes appear to have been confined to the Northern Hemisphere, having been found in North America, Asia, and Europe, and possibly also North Africa. Choristoderes are generally thought to be derived neodiapsids that are close relatives or members of Sauria.
| Choristodera | |
|---|---|

| |
| Skeleton of Philydrosaurus proselius exhibited at the National Museum of Natural Science, Taiwan | |
.jpg)
| |
| Skeleton of Ikechosaurus, on display at National Museum of Natural Science, Taiwan | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Clade: | Neodiapsida |
| Order: | †Choristodera Cope, 1876 |
| Subgroups | |
| |
History of discovery
edit
Choristodera was erected in 1876, originally as a suborder of Rhynchocephalia by Edward Drinker Cope to contain Champsosaurus, which was described from Late Cretaceous strata of Montana by Cope in the same paper.[3][4] A year later, in 1877, Simoedosaurus was described by Paul Gervais from Upper Paleocene deposits at Cernay, near Rheims, France. These remained the only recognised choristoderes for over a century, until new taxa were described in the late 20th century.[5] Beginning in the late 1970s, additional taxa were described by Soviet-Mongolian teams from Lower Cretaceous sediments in Mongolia. In studies from 1989 to 1991, Susan E. Evans described new material of Cteniogenys from the Middle Jurassic of Britain. The genus had first been described by Charles W. Gilmore in 1928 from the Late Jurassic of the western United States, and had previously been enigmatic. The studies revealed it to be a small, lizard-like choristodere, different from the crocodile-like forms previously known.[6]
Description
edit_NMNS.jpg)
Choristoderes vary substantially in size, the smallest genera like Cteniogenys and Lazarussuchus had a length of only around 30 cm (12 in), and the largest known choristoderan, Kosmodraco dakotensis is estimated to have had a total length of around 5 m (16 ft).[5][1] Neochoristoderes such as Champsosaurus are the best-known group of the Choristodera. They resembled modern crocodilians, especially gharials. The skull of these animals have a long, thin snout filled with small, sharp conical teeth. Other choristoderes are referred to collectively as "non-neochoristoderes", which are mostly small lizard-like forms, though Shokawa, Khurendukhosaurus and Hyphalosaurus possess long plesiosaur like necks. The grouping of "non-neochoristoderes" is paraphyletic (not containing all descendants of a common ancestor), as the lizard-like bodyform represents the ancestral morphology of the group.[5]
Skeletal anatomy
edit
According to Matsumoto and colleagues (2019), choristoderes are united by the presence of nine synapomorphies (shared traits characteristic of the group), including a median contact of the elongated prefrontal bones of the skull separating the nasal bones from the frontal bones, the dorsal flange of the maxilla is inflected medially (toward the midline of the body), the parietal foramen are absent, the squamosal bones are expanded behind (posterior to) the occipital condyle, the teeth are conical and sub-thecodont (located in shallow sockets), the dentaries are slender with elongated grooves running along the labial (outward facing) surface of the bone, additional sacral vertebrae are present, expanded "spine tables" are present on the vertebrae, and the surfaces of both ends of vertebral centra are flat (amphiplatyan).[7] All known choristoderans possess or are inferred to possess a novel skull bone not found in other reptiles, referred to as the "neomorphic bone" or neomorph, which is a component of the dermatocranium.[8][9] Ancestrally, the skull of choristoderes possess elongated upper and lower temporal fenestrae (openings of the skull behind the eye socket), these are greatly expanded in neochoristoderes, most extremely in Champsosaurus, giving the skull a cordiform (heart shaped) appearance when viewed from above.[8] In many "non-neochoristoderes" the lower temporal fenestrae are secondarily closed.[10] Choristoderes possessed gastralia (rib-like bones situated in the abdomen) like tuatara and crocodilians.[11]
-
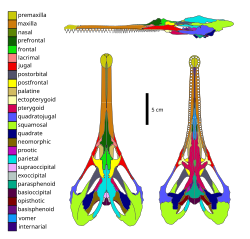 Skull diagram of Champsosaurus lindoei, a neochoristoderan
Skull diagram of Champsosaurus lindoei, a neochoristoderan -
 Skull of Lazarussuchus, an allochoristodere
Skull of Lazarussuchus, an allochoristodere


Internal skull anatomy
editThe internal skull anatomy of choristoderes is only known for Champsosaurus. The braincase of Champsosaurus is poorly ossified at the front of the skull (anterior), but is well ossified in the rear (posterior) similar to other diapsids. The cranial endocast (space occupied by the brain in the cranial vault) is proportionally narrow in both lateral and dorsoventral axes, with an enlarged pineal body and olfactory bulbs. The optic lobes and flocculi are small in size, indicating only average vision ability at best. The olfactory chambers of the nasal passages and olfactory stalks of the braincase are reasonably large, indicating that Champsosaurus probably had good olfactory capabilities (sense of smell). The nasal passages lack bony turbinates. The semicircular canals of the inner ear are most similar to those of other aquatic reptiles. The expansion of the sacculus indicates that Champsosaurus likely had an increased sensitivity to low frequency sounds and vibrations.[12]
Dentition
editMost choristoderes have rather simple undifferentiated (homodont) teeth, with striated enamel covering the tooth crown but not the base. Neochoristoderes have teeth completely enveloped in striated enamel with an enamel infolding at the base, labiolingually compressed and hooked, the exception being Ikechosaurus which has still rather simple teeth aside from the start of an enamel infolding. Teeth implantation is subthecodont, with teeth being replaced by erosion of a pit in the lingual (side of the tooth facing the tongue) surface of the tooth base. There is some tooth differentiation among neochoristoderes, with the anterior teeth being sharper and more slender than posterior teeth. Choristoderes retain palatal teeth (teeth present on the bones of the roof of the mouth). Unlike most diapsid groups, where palatal teeth are reduced or lost completely, the palatal teeth in choristoderes are extensively developed indicating food manipulation in the mouth, probably in combination with the tongue. In most choristoderes, longitudinal rows of palatal teeth are present on the pterygoid, palatine and vomer, as well as a row on the pterygoid flange. In some neochoristoderes the palatal tooth rows are modified into tooth batteries on raised platforms. The morphology of the palatal teeth is identical to that of the marginal teeth of non-neochoristoderes, and the replacement of palatal teeth is nearly identical to the replacement of marginal teeth.[13]
Skin
edit

An exceptionally preserved specimen of Monjurosuchus preserves pleated skin, which indicates that in life it was probably thin and soft. The preserved scales are small and overlapping, and are smaller on the ventral underside of the body than the dorsal surface. A double row of larger ovoid scales runs along the dorsum (upper midline) of the body. The fossil also preserves webbed feet.[14] Hyphalosaurus was covered in scales of varying shape, depending on their position on the body, with at least one and possibly multiple rows of large ovoid scales running down sides of the trunk and tail. The feet display evidence of webbing, and the tail probably had additional tissue at the top and bottom, allowing it to be used as a fin to propel Hyphalosaurus through the water.[11][15] Skin impressions of Champsosaurus have also been reported, they consist of small (0.6-0.1 mm) pustulate and rhomboid scales, with the largest scales being located on the lateral sides of the body, decreasing in size dorsally, no osteoderms were present.[16] The Menat specimen of Lazarussuchus preserves some remnants of soft tissue, but no scales, which shows that the hindfoot (pes) was not webbed, and a dark stained region with a crenellated edge is present above the caudal vertebrae of the tail, suggestive of a crest similar to those found in some living reptiles, like the tuatara, lizards and crocodiles.[17]
Paleobiology
editChoristoderes are exclusively found in freshwater deposits, often associated with turtles, fish, frogs, salamanders and crocodyliformes. They appear to have been almost exclusively found in warm temperate climates, with the range of neochoristoderes extending to the high Canadian Arctic during the Coniacian-Santonian stages of the Late Cretaceous (~89-83 Million years ago), a time of extreme warmth. Due to the morphological similarities between choristoderes and crocodyliformes, it has often been assumed that they existed in competition. However "non-neochoristoderes" were smaller than adult aquatic crocodyliformes and were more likely in competition with other taxa. For the more crocodile-like neochoristoderes, there appears to have been niche differentiation, with gharial-like neochoristoderans occurring in association with blunt snouted crocodyliformes, but not in association with narrow snouted forms.[5]
Diet
editNeochoristoderans are presumed to have been piscivorous.[16] Champsosaurus in particular is thought to have fed like modern gharials, sweeping its head to the side to catch individual fish from shoals, while Simoedosaurus is thought to have been more generalist, being able to take both aquatic and terrestrial prey.[18] Cteniogenys and Lazarussuchus have been suggested to have fed on invertebrates.[6] Preserved gut contents of a Monjurosuchus specimen appear to show arthropod cuticle fragments.[14] Another specimen of Monjurosuchus has been found with preserved skulls of seven juvenile individuals within the abdominal cavity. This has been proposed to represent evidence of cannibalism.[19] However, this proposal has been criticised by other authors, who suggest it is more likely that they represent late-stage embryos.[20] A specimen of Hyphalosaurus has been found with small rib bones in its abdominal cavity, suggesting that it took vertebrate prey at least on occasion.[11]
Reproduction
editA specimen of Hyphalosaurus baitaigouensis has been found with 18 fully developed embryos within the mother's body, suggesting that they were viviparous,[21] but another specimen shows that Hyphalosaurus baitaigouensis also possessed soft-shelled eggs, similar to those of lepidosaurs.[22] A possible explanation for this is that Hyphalosaurus was ovoviviparous, with the thin-shelled eggs hatching immediately after they were laid, presumably on land,[23] though it has also been suggested that the species employed both viviparous and oviparous reproductive modes.[20] An embryo of Ikechosaurus has been found preserved within a weakly mineralised parchment-shelled egg, suggesting that Ikechosaurus was oviparous, and laid their eggs on land.[20] Monjuruosuchus has been suggested to have been viviparous.[20] In Champsosaurus, it has been suggested that adult females could crawl ashore to lay eggs on land, with males and juveniles appearing to be incapable of doing so, based on the presumably sexually dimorphic fusion of the sacral vertebrae and possession of more robust limb bones in presumed females.[24] A skeleton of Philydrosaurus has been found with associated post-hatchling stage juveniles, suggesting that they engaged in post-hatching parental care.[23]
Tracks
editTracks from the Early Cretaceous (Albian) of South Korea, given the ichnotaxon name Novapes ulsanensis have been attributed to choristoderans, based on the similarity of the pentadactyl (five fingered) preserved tracks to the foot morphology of Monjurosuchus. The tracks preserve traces of webbing between the digits. The authors of the study proposed based on the spacing of the prints, that choristoderans could "high walk" like modern crocodilians.[25] Tracks attributed to neochoristoderans dubbed Champsosaurichnus parfeti have also been reported from the Late Cretaceous Laramie Formation of the United States, though only two prints are present and it is not possible to distinguish between a manus (forefoot) or pes (hindfoot).[26]
Classification and phylogeny
edit
Internal systematics
editHistorically, the internal phylogenetics of Choristodera were unclear, with the neochoristoderes being recovered as a well-supported clade, but the relationships of the "non-neochoristoderes" being poorly resolved.[7] However, during the 2010s, the "non-neochoristoderes" from the Early Cretaceous of Asia (with the exception of Heishanosaurus) alongside Lazarussuchus from the Cenozoic of Europe were recovered (with weak support) as belonging to a monophyletic clade, which were informally named the "Allochoristoderes" by Dong and colleagues in 2020, characterised by the shared trait of completely closed lower temporal fenestrae, with Cteniogenys from the Middle-Late Jurassic of Europe and North America being consistently recovered as the basalmost choristodere.[10] The long necked "non-neochoristoderes" Shokawa and Hyphalosaurus have often been recovered as a clade, dubbed the Hyphalosauridae by Gao and Fox in 2005.[27] The finding of more complete material of the previously fragmentary Khurendukhosaurus shows that it also has a long neck, and it has also been recovered as part of the clade.[28]
Phylogeny from the analysis of Dong and colleagues (2020):[10]
| Choristodera |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Relationships to other reptiles
editChoristoderes are universally agreed to be members of Neodiapsida, but their exact placement in the clade is uncertain, due to their mix of primitive and derived features, and a long ghost lineage (absence of a fossil record) after their split from other reptiles.[29] After initially being placed in Rhynchocephalia, Cope later suggested a placement in Lacertilla due to the shape of the cervical vertebrae. Louis Dollo in 1891 returned Choristodera to Rhynchocephalia, but in 1893 suggested a close relationship with Pareiasaurus. Alfred Romer in publications in 1956 and 1968 placed Choristodera within the paraphyletic or polyphyletic grouping of "Eosuchia", describing them, as “an offshoot of the basic eosuchian stock”, a classification which was widely accepted. However, the use of computer based cladistics in the 1980s demonstrated the non-monophyly of "Eosuchia", making the classification of choristoderes again uncertain.[30] Subsequent studies either suggested placement as archosauromorphs, lepidosauromorphs or members of Diapsida incertae sedis. In a 2016 analysis of neodiapsid relationships by Martín Ezcurra they were recovered as members of the advanced neodiapsid group Sauria, in a polytomy with Lepidosauromorpha and Archosauromorpha, with being the earliest diverging members of either group also being plausible.[29] A position as basal archosauromorphs is supported by the ossification sequence of their embryos.[20]
Evolutionary history
edit
Choristoderes must have diverged from all other known reptile groups prior to the end of the Permian period, over 250 million years ago, based on their primitive phylogenetic position.[5] In 2015, Rainer R. Schoch reported a new small (~ 20 cm long) diapsid from the Middle Triassic (Ladinian) Lower Keuper of Southern Germany, known from both cranial and postcranial material, which he claimed represented the oldest known choristodere.[31] Pachystropheus from the Late Triassic (Rhaetian) of Britain was historically suggested to be a choristodere,[32] but was later demonstrated to be a member of the marine reptile group Thalattosauria.[33] The oldest unequivocal choristoderan is the small lizard-like Cteniogenys, the oldest known remains of which are known from the late Middle Jurassic (Bathonian ~168-166 million years ago) Forest Marble and Kilmaluag formations of Britain, with remains also known from the Upper Jurassic Alcobaça Formation of Portugal and the Morrison Formation of the United States, with broadly similar remains also known from the late Middle Jurassic (Callovian) Balabansai Formation of Kyrgyzstan in Central Asia,[5][34] the Bathonian Itat Formation of western Siberia,[35] as well as possibly the Bathonian aged Anoual Formation in Morocco, North Africa.[36]
Choristoderes underwent a major evolutionary radiation in Asia during the Early Cretaceous, which represents the high point of choristoderan diversity, including the first records of the gharial-like Neochoristodera, which appear to have evolved in the regional absence of aquatic neosuchian crocodyliformes.[5] A partial femur of an indeterminate choristodere is known from the Yellow Cat Member of the Cedar Mountain Formation in North America.[37] They appear to be absent from the well sampled European localities of the Berriasian aged Purbeck Group, Great Britain and the Barremian aged La Huérguina Formation, Spain,[5] though there is a record of a small Cteniogenys-like taxon from the Berriasian aged Angeac-Charente bonebed in France.[38] In the latter half of the Late Cretaceous (Campanian-Maastrichtian), the neochoristodere Champsosaurus is found in Utah, Wyoming, Montana, North Dakota, Alberta and Saskatchewan, which were along the western coast of the Western Interior Seaway on the island of Laramidia.[5] Indeterminate remains of neochoristoderes are also known from the Canadian High Arctic, dating to the early Late Cretaceous (Coniacian–Turonian)[39] and from the Navesink Formation of New Jersey from the latest Cretaceous (Maastrichtian), which formed the separate island of Appalachia.[40] Vertebrae from the Cenomanian of Germany[41] and the Campanian aged Grünbach Formation of Austria[42] indicate the presence of choristoderes in Europe during this time period. The only record of choristoderes from Asia in the Late Cretaceous is a single vertebra from the Turonian of Japan.[43] Fragmentary remains found in the Campanian aged Oldman and Dinosaur Park formations in Alberta, Canada, also possibly suggest the presence of small bodied "non-neochoristoderes" in North America during the Late Cretaceous.
Champsosaurus survived the K-Pg extinction, and together with fellow neochoristodere Simoedosaurus are present in Europe, Asia and North America during the Paleocene, however they became extinct during the early Eocene. Their extinction coincides with major faunal turnover associated with elevated temperatures.[5] Small bodied "non-neochoristoderes", which are absent from the fossil record after the Early Cretaceous (except for possible North American remains), reappear in the form of the lizard-like Lazarussuchus from the late Paleocene of France.[17] The European endemic Lazarussuchus is the last known choristodere, surviving the extinction of neochoristoderes at the beginning of the Eocene, with the youngest known remains being those of L. dvoraki from the Early Miocene of the Czech Republic[5][44] and as well as possible indeterminate remains of Lazarussuchus reported from the late Miocene (~11.6 million years ago) of southern Germany.[45]
References
edit- ^ a b Brownstein, C. D. (2022). "High morphological disparity in a bizarre Paleocene fauna of predatory freshwater reptiles". BMC Ecology and Evolution. 22 (1): 34. doi:10.1186/s12862-022-01985-z. PMC 8935759. PMID 35313822.
- ^ Sues, H.-D. (2019). "Archosauromorpha: Choristodera". The Rise of Reptiles: 320 Million Years of Evolution. Baltimore, MD: Johns Hopkins University Press. pp. 167–168. ISBN 978-1-4214-2867-3.
- ^ Cope ED (1876). "On some extinct reptiles and Batrachia from the Judith River and Fox Hills beds of Montana". Proceedings of the Academy of Natural Sciences of Philadelphia. 1876: 340–359.
- ^ Cope, ED (1884). "The Choristodera". The American Naturalist. 18: 815–817.
- ^ a b c d e f g h i j k Matsumoto R, Evans SE (2010). "Choristoderes and the freshwater assemblages of Laurasia". Journal of Iberian Geology. 36 (2): 253–274. Bibcode:2010JIbG...36..253M. doi:10.5209/rev_jige.2010.v36.n2.11.
- ^ a b Evans SE, Hecht MK (1993). "A History of an Extinct Reptilian Clade, the Choristodera: Longevity, Lazarus-Taxa, and the Fossil Record". In Hecht M, MacIntyre RJ, Clegg MT (eds.). Evolutionary Biology. Boston, MA: Springer US. pp. 323–338. doi:10.1007/978-1-4615-2878-4_8. ISBN 978-1-4615-2878-4.
- ^ a b Matsumoto R, Dong L, Wang Y, Evans SE (2019). "The first record of a nearly complete choristodere (Reptilia: Diapsida) from the Upper Jurassic of Hebei province, People's Republic of China" (PDF). Journal of Systematic Palaeontology. 17 (12): 1031–1048. Bibcode:2019JSPal..17.1031M. doi:10.1080/14772019.2018.1494220. S2CID 92421503.
- ^ a b Dudgeon TW, Maddin HC, Evans DC, Mallon JC (April 2020). "Computed tomography analysis of the cranium of Champsosaurus lindoei and implications for the choristoderan neomorphic ossification". Journal of Anatomy. 236 (4): 630–659. doi:10.1111/joa.13134. PMC 7083570. PMID 31905243.
- ^ Qin, Wanying; Yi, Hongyu; Gao, Keqin (2 July 2022). "A neomorphic ossification connecting the braincase, squamosal, and quadrate in choristoderan reptiles: insights from µCT data". Fossil Record. 25 (1): 1–10. Bibcode:2022FossR..25....1Q. doi:10.3897/fr.25.79595. ISSN 2193-0074.
- ^ a b c Dong, Liping; Matsumoto, Ryoko; Kusuhashi, Nao; Wang, Yuanqing; Wang, Yuan; Evans, Susan E. (2 August 2020). "A new choristodere (Reptilia: Choristodera) from an Aptian–Albian coal deposit in China". Journal of Systematic Palaeontology. 18 (15): 1223–1242. Bibcode:2020JSPal..18.1223D. doi:10.1080/14772019.2020.1749147. ISSN 1477-2019. S2CID 219047160.
- ^ a b c Gao KQ, Ksepka DT (June 2008). "Osteology and taxonomic revision of Hyphalosaurus (Diapsida: Choristodera) from the Lower Cretaceous of Liaoning, China". Journal of Anatomy. 212 (6): 747–68. doi:10.1111/j.1469-7580.2008.00907.x. PMC 2423398. PMID 18510504.
- ^ Dudgeon, Thomas W.; Maddin, Hillary C.; Evans, David C.; Mallon, Jordan C. (28 April 2020). "The internal cranial anatomy of Champsosaurus (Choristodera: Champsosauridae): Implications for neurosensory function". Scientific Reports. 10 (1): 7122. Bibcode:2020NatSR..10.7122D. doi:10.1038/s41598-020-63956-y. ISSN 2045-2322. PMC 7188685. PMID 32346021.
- ^ Matsumoto R, Evans SE (March 2016). "Morphology and function of the palatal dentition in Choristodera". Journal of Anatomy. 228 (3): 414–29. doi:10.1111/joa.12414. PMC 5341546. PMID 26573112.
- ^ a b Keqin G, Evans S, Qiang J, Norell M, Shu'An J (25 September 2000). "Exceptional fossil material of a semi-aquatic reptile from China: the resolution of an enigma". Journal of Vertebrate Paleontology. 20 (3): 417–421. doi:10.1671/0272-4634(2000)020[0417:EFMOAS]2.0.CO;2. S2CID 86064702.
- ^ Wang, Miaoyan; Xing, Lida; Niu, Kecheng; Liang, Qingqing; Evans, Susan E. (April 2023). "A new specimen of the Early Cretaceous long-necked choristodere Hyphalosaurus from Liaoning, China with exceptionally-preserved integument". Cretaceous Research. 144: 105451. Bibcode:2023CrRes.14405451W. doi:10.1016/j.cretres.2022.105451.
- ^ a b Erickson BR (June 1985). "Aspects of some anatomical structures of Champsosaurus (Reptilia: Eosuchia)". Journal of Vertebrate Paleontology. 5 (2): 111–127. Bibcode:1985JVPal...5..111E. doi:10.1080/02724634.1985.10011849.
- ^ a b Matsumoto R, Buffetaut E, Escuillie F, Hervet S, Evans SE (2013). "New material of the choristodere Lazarussuchus (Diapsida, Choristodera) from the Paleocene of France". Journal of Vertebrate Paleontology. 33 (2): 319–339. Bibcode:2013JVPal..33..319M. doi:10.1080/02724634.2012.716274. S2CID 129438118.
- ^ Matsumoto, Ryoko; Fujiwara, Shin-ichi; Evans, Susan E. (2021). "Feeding behaviour and functional morphology of the neck in the long-snouted aquatic fossil reptile Champsosaurus (Reptilia: Diapsida) in comparison with the modern crocodilian Gavialis gangeticus". Journal of Anatomy. 240 (5): 893–913. doi:10.1111/joa.13600. ISSN 1469-7580. PMC 9005684. PMID 34865223. S2CID 244917425.
- ^ Wang, Xiaolin; Miao, Desui; Zhang, Yuguang (1 February 2005). "Cannibalism in a semi-aquatic reptile from the Early Cretaceous of China". Chinese Science Bulletin. 50 (3): 282–284. Bibcode:2005ChSBu..50..282W. doi:10.1007/BF02897540. ISSN 1861-9541. S2CID 95194181.
- ^ a b c d e Jiang, Baoyu; He, Yiming; Elsler, Armin; Wang, Shengyu; Keating, Joseph N.; Song, Junyi; Kearns, Stuart L.; Benton, Michael J. (12 June 2023). "Extended embryo retention and viviparity in the first amniotes". Nature Ecology & Evolution. 7 (7): 1131–1140. Bibcode:2023NatEE...7.1131J. doi:10.1038/s41559-023-02074-0. ISSN 2397-334X. PMC 10333127. PMID 37308704.
- ^ Ji Q, Wu XC, Cheng YN (April 2010). "Cretaceous choristoderan reptiles gave birth to live young". Die Naturwissenschaften. 97 (4): 423–8. Bibcode:2010NW.....97..423J. doi:10.1007/s00114-010-0654-2. PMID 20179895. S2CID 8719805.
- ^ Hou LH, Li PP, Ksepka DT, Gao KQ, Norell MA (April 2010). "Implications of flexible-shelled eggs in a Cretaceous choristoderan reptile". Proceedings. Biological Sciences. 277 (1685): 1235–9. doi:10.1098/rspb.2009.2035. PMC 2842823. PMID 20018793.
- ^ a b Lü J, Kobayashi Y, Deeming DC, Liu Y (20 October 2014). "Post-natal parental care in a Cretaceous diapsid from northeastern China" (PDF). Geosciences Journal. 19 (2): 273–280. doi:10.1007/s12303-014-0047-1. S2CID 49570935.
- ^ Katsura Y (2007). "Fusion of sacrals and anatomy in Champsosaurus(Diapsida, Choristodera)". Historical Biology. 19 (3): 263–271. Bibcode:2007HBio...19..263K. doi:10.1080/08912960701374659. S2CID 84966652.
- ^ Lee YN, Kong DY, Jung SH (September 2020). "The first possible choristoderan trackway from the Lower Cretaceous Daegu Formation of South Korea and its implications on choristoderan locomotion". Scientific Reports. 10 (1): 14442. Bibcode:2020NatSR..1014442L. doi:10.1038/s41598-020-71384-1. PMC 7468130. PMID 32879388.
- ^ Lockley, Martin G.; Hunt, Adrian P. (14 September 1995). "Ceratopsid tracks and associated ichnofauna from the Laramie Formation (Upper Cretaceous: Maastrichtian) of Colorado". Journal of Vertebrate Paleontology. 15 (3): 592–614. Bibcode:1995JVPal..15..592L. doi:10.1080/02724634.1995.10011251. ISSN 0272-4634.
- ^ Gao, Ke-Qin; Fox, Richard C. (16 November 2005). "A new choristodere (Reptilia: Diapsida) from the Lower Cretaceous of western Liaoning Province, China, and phylogenetic relationships of Monjurosuchidae". Zoological Journal of the Linnean Society. 145 (3): 427–444. doi:10.1111/j.1096-3642.2005.00191.x. ISSN 1096-3642.
- ^ Matsumoto, Ryoko; Tsogtbaatar, Khishigjav; Ishigaki, Shinobu; Tsogtbaatar, Chinzorig; Enkhtaivan, Zorig; Evans, Susan (2019). "Revealing body proportions of the enigmatic choristodere Khurendukhosaurus from Mongolia". Acta Palaeontologica Polonica. 64. doi:10.4202/app.00561.2018. S2CID 133925482.
- ^ a b Ezcurra MD (28 April 2016). "The phylogenetic relationships of basal archosauromorphs, with an emphasis on the systematics of proterosuchian archosauriforms". PeerJ. 4: e1778. doi:10.7717/peerj.1778. ISSN 2167-8359. PMC 4860341. PMID 27162705.
- ^ Gao, Keqin; Fox, Richard C. (December 1998). "New choristoderes (Reptilia: Diapsida) from the Upper Cretaceous and Palaeocene, Alberta and Saskatchewan, Canada, and phylogenetic relationships of Choristodera". Zoological Journal of the Linnean Society. 124 (4): 303–353. doi:10.1111/j.1096-3642.1998.tb00580.x.
- ^ Schoch, R.R (2015). "Reptilien" (PDF). Der Lettenkeuper: ein Fenster in die Zeit vor den Dinosauriern (in German). Staatliches Museum für Naturkunde Stuttgart. pp. 231–264.
- ^ Storrs, G. W.; Gower, D. J. (1 November 1993). "The earliest possible choristodere (Diapsida) and gaps in the fossil record of semi-aquatic Reptiles". Journal of the Geological Society. 150 (6): 1103–1107. Bibcode:1993JGSoc.150.1103S. doi:10.1144/gsjgs.150.6.1103. ISSN 0016-7649. S2CID 86088809.
- ^ Quinn, Jacob G.; Matheau-Raven, Evangelos R.; Whiteside, David I.; Marshall, John E. A.; Hutchinson, Deborah J.; Benton, Michael J. (4 June 2024). "The relationships and paleoecology of Pachystropheus rhaeticus , an enigmatic latest Triassic marine reptile (Diapsida: Thalattosauria)". Journal of Vertebrate Paleontology. doi:10.1080/02724634.2024.2350408. ISSN 0272-4634.
- ^ Averianov, Alexander O.; Martin, Thomas; Evans, Susan E.; Bakirov, Aizek A. (1 January 2006). "First Jurassic Choristodera from Asia". Naturwissenschaften. 93 (1): 46–50. Bibcode:2006NW.....93...46A. doi:10.1007/s00114-005-0061-2. ISSN 1432-1904. PMID 16344982. S2CID 7111980.
- ^ "Middle Jurassic vertebrate assemblage of Berezovsk coal mine in western Siberia (Russia)". Global Geology. 19 (4): 187–204. 2016. doi:10.3969/j.issn.1673-9736.2016.04.01.
- ^ Haddoumi H, Allain R, Meslouh S, Metais G, Monbaron M, Pons D, et al. (2016). "Guelb el Ahmar (Bathonian, Anoual Syncline, eastern Morocco): First continental flora and fauna including mammals from the Middle Jurassic of Africa" (PDF). Gondwana Research. 29 (1): 290–319. Bibcode:2016GondR..29..290H. doi:10.1016/j.gr.2014.12.004.
- ^ Britt, Brooks B.; Scheetz, Rodney D.; Brinkman, Donald B.; Eberth, David A. (11 December 2006). "A Barremian neochoristodere from the Cedar Mountain Formation, Utah, U.S.A." Journal of Vertebrate Paleontology. 26 (4): 1005–1008. doi:10.1671/0272-4634(2006)26[1005:abnftc]2.0.co;2. ISSN 0272-4634. S2CID 86258448.
- ^ Ronan Allain, Romain Vullo, Lee Rozada, Jérémy Anquetin, Renaud Bourgeais, et al.. Vertebrate paleobiodiversity of the Early Cretaceous (Berriasian) Angeac-Charente Lagerstätte (southwestern France): implications for continental faunal turnover at the J/K boundary. Geodiversitas, Museum National d’Histoire Naturelle Paris, In press. ffhal-03264773f
- ^ Vandermark, Deborah; Tarduno, John A.; Brinkman, Donald B. (May 2007). "A fossil champsosaur population from the high Arctic: Implications for Late Cretaceous paleotemperatures". Palaeogeography, Palaeoclimatology, Palaeoecology. 248 (1–2): 49–59. Bibcode:2007PPP...248...49V. doi:10.1016/j.palaeo.2006.11.008.
- ^ Dudgeon, Thomas W.; Landry, Zoe; Callahan, Wayne R.; Mehling, Carl M.; Ballwanz, Steven (2021). "An Appalachian population of neochoristoderes (Diapsida, Choristodera) elucidated using fossil evidence and ecological niche modelling". Palaeontology. 64 (5): 629–643. Bibcode:2021Palgy..64..629D. doi:10.1111/pala.12545. ISSN 1475-4983. S2CID 237761128.
- ^ Reiss S, Scheer U, Sachs S, Kear BP (13 December 2018). "Filling the biostratigraphical gap: First choristoderan from the Lower - mid-Cretaceous interval of Europe". Cretaceous Research. 96: 135–141. doi:10.1016/j.cretres.2018.12.009. ISSN 0195-6671. S2CID 134904339.
- ^ Buffetaut, Eric (1989). "Erster nachweis von Choristodera (Reptilia, Diapsida) in der Oberkreide Europas: Champsosaurierwirbel aus den Gosau-Schichten (Campan) Niederösterreichs" [First evidence of Choristodera (Reptilia, Diapsida) in the Upper Cretaceous of Europe: Champsosaur vertebrae from the Gosau strata (Campanian) of Lower Austria] (PDF). Sitzungsberichten der Österreichs Akademis der Wissenschaften Mathematisch-Naturwissenschaftlichen Klasse, Abteilung (in German). 197: 389–394.
- ^ Matsumoto, Ryoko; Hirayama, Ren; Miyata, Shinya; Yoshida, Masataka; Mitsuzuka, Shunsuke; Takisawa, Toshio; Evans, Susan E. (August 2021). "The first choristoderan record from the Upper Cretaceous of Asia, Tamagawa Formation, Kuji Group, Japan". Cretaceous Research. 129: 104999. doi:10.1016/j.cretres.2021.104999. ISSN 0195-6671. S2CID 238680387.
- ^ Evans SE, Klembara J (2005). "A choristoderan reptile (Reptilia: Diapsida) from the Lower Miocene of northwest Bohemia (Czech Republic)". Journal of Vertebrate Paleontology. 25 (1): 171–184. doi:10.1671/0272-4634(2005)025[0171:ACRRDF]2.0.CO;2. ISSN 0272-4634. S2CID 84097919.
- ^ Kirscher, U.; Prieto, J.; Bachtadse, V.; Aziz, H. Abdul; Doppler, G.; Hagmaier, M.; Böhme, M. (1 August 2016). "A biochronologic tie-point for the base of the Tortonian stage in European terrestrial settings: Magnetostratigraphy of the topmost Upper Freshwater Molasse sediments of the North Alpine Foreland Basin in Bavaria (Germany)" (PDF). Newsletters on Stratigraphy. 49 (3): 445–467. doi:10.1127/nos/2016/0288. ISSN 0078-0421.
Further reading
edit- de Braga M, Rieppel O (1997). "Reptile phylogeny and the interrelationships of turtles". Zoological Journal of the Linnean Society. 120 (3): 281–354. doi:10.1111/j.1096-3642.1997.tb01280.x.
- Erickson BR (1972). The Lepidosaurian Reptile Champsosaurus in North America. Paleontology, Monograph. Vol. 1. Science Museum of Minnesota.
- Matsumoto R, Suzuki S, Tsogtbaatar K, Evans SE (February 2009). "New material of the enigmatic reptile Khurendukhosaurus (Diapsida: Choristodera) from Mongolia". Die Naturwissenschaften. 96 (2): 233–42. Bibcode:2009NW.....96..233M. doi:10.1007/s00114-008-0469-6. PMID 19034405. S2CID 13542692.
- Ksepka D, Gao K, Norell MA (2005). "A new choristodere from the Cretaceous of Mongolia". American Museum Novitates (3468): 1–22. doi:10.1206/0003-0082(2005)468<0001:ancftc>2.0.co;2. hdl:2246/2778. S2CID 54179348.