
An Error has occurred retrieving Wikidata item for infobox
Terminal deoxynucleotidyl transferase (TdT), also known as DNA nucleotidylexotransferase (DNTT) or terminal transferase, is a specialized DNA polymerase expressed in immature, pre-B, pre-T lymphoid cells, and acute lymphoblastic leukemia/lymphoma cells. TdT adds N-nucleotides to the V, D, and J exons of the TCR and BCR genes during antibody gene recombination, enabling the phenomenon of junctional diversity. In humans, terminal transferase is encoded by the DNTT gene.[1][2] As a member of the X family of DNA polymerase enzymes, it works in conjunction with polymerase λ and polymerase μ, both of which belong to the same X family of polymerase enzymes. The diversity introduced by TdT has played an important role in the evolution of the vertebrate immune system, significantly increasing the variety of antigen receptors that a cell is equipped with to fight pathogens. Studies using TdT knockout mice have found drastic reductions (10-fold) in T-cell receptor (TCR) diversity compared with that of normal, or wild-type, systems. The greater diversity of TCRs that an organism is equipped with leads to greater resistance to infection.[3][4] Althought TdT was one of the first DNA polymerases identified in mammals in 1960[5], it remains one of the least understood of all DNA catalysis enzymes[6]. In 2016-18, TdT was discovered to demonstrate in trans template dependant behaviour in addition to its more broadly known template independant behaviour[7][8].
TdT is absent in fetal liver HSCs, significantly impairing junctional diversity in B-cells during the fetal period.[9]
Function and regulation
editGenerally, TdT catalyses the addition of nucleotides to the 3' terminus of a DNA molecule. Unlike most DNA polymerases, it does not require a template. The preferred substrate of this enzyme is a 3'-overhang, but it can also add nucleotides to blunt or recessed 3' ends. Futher, TdT is the only polymerase that is know to catalyze the synthesis of 2-15nt DNA polymers from free nucleotides in solution in vivo[10]. In vitro, this behaviour catalyzes the general formation of DNA polymers without specific length[11]. The 2-15nt DNA fragments produced in vivo are hypothesized to act in signaling pathways related to DNA repair and/or recombination machinery[10]. Like many polymerases, TdT requires a divalent cation cofactor[12], however, TdT is unique in it's ability to use a broader range of cations such as Mg2+, Mn2+, Zn2+ and Co2+[12]. The rate of enzymatic activity depends on the available divalent cations and the nucleotide being added. [13]
TdT is expressed mostly in the primary lymphoid organs, like the thymus and bone marrow. Regulation of its expression occurs via multiple pathways. These include protein-protein interactions, like those with TdIF1. TdIF1 is another protein that interacts with TdT to inhibit its function by masking the DNA binding region of the TdT polymerase. The regulation of TdT expression also exists at the transcriptional level, with regulation influenced by stage-specific factors, and occurs in a developmentally restrictive manner.[3][14][15] Although expression is typically found to be in the primary lymphoid organs, recent work has suggested that stimulation via antigen can result in secondary TdT expression along with other enzymes needed for gene rearrangement outside of the thymus for T-cells.[16] Patients with acute lymphoblastic leukemia greatly over-produce TdT[13]. Cell lines derived from these patients served as one of the first sources of pure TdT and lead to the discovery that differences in activity exist between human and bovine isoforms[13].
Mechanism
edit
Similar to many polymerases, the catalytic site of TdT has two divalent cations in its palm domain that assist in nucleotide binding, help lower the pKa of the 3'-OH group and ultimately facilitate the departure of the resultant pyrophosphate by-product[17][18].
Isoform Variation
editSeveral isoforms of TdT have observed in mice, bovines, and humans. To date, two varients have been identified in mice while three have been identified in humans[19].
The two splice variants identified in mice are named according to their respective lengths: TdTS consists of 509 amino acids while TdTL, the longer variant, consists of 529 amino acids. The differences between TdTS and TdTL occur outside regions that bind DNA and nucleotides. That the 20 amino acid difference has an effect on enzymatic activity is controversial, with some arguing that TdTL's modifications bestow exonuclease activity while others argue that TdTL and TdTS have nearly identical in vitro activity. Additionally, TdTL reportedly can modulate the catalytic activity of TdTS in vivo through an unknown mechanism. It is suggested that this aids in the regulation of TdT's role in V(D)J recombination[20].
Human TdT isoforms have three varients TdTL1, TdTL2, and TdTS. TdTL1 is broadly expressed in lymphoid cell lines while TdTL2 is predominantly expressed in normal small lymphocytes. Both localize in the nucleus when expressed[21] and both possess 3'->5' exonuclease activity[22]. In contrast, TdTS isoforms do not possess exonuclease activity and perform the necessary elongation during V(D)J recombination[22]. Since a similar exonuclease activity hypothesized in murine TdTL is found in human and bovine TdTL which has lead some to postulate that that bovine and human TdTL isoforms regulate TdTS isoforms in a similar manner as proposed in mice[20]. Further, some hypothesize that TdTL1 may be involved in the regulation of TdTL2 and/or TdTS activity.
Role in V(D)J Recombination
edit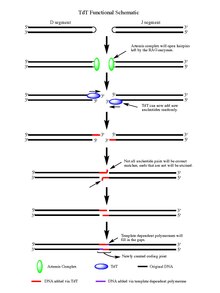
Upon the action of the RAG 1/2 enzymes, the cleaved double stranded DNA is left with hairpin structures at the end of each DNA segment created by the cleavage event. The hairpins are both opened by the Artemis complex, which has endonuclease activity when phosphorylated, providing the free 3' OH ends for TdT to act upon. Once the Artemis complex has done its job and added palindromic nucleotides (P-nucleotides) to the newly opened DNA hairpins, the stage is set for TdT to do its job. TdT is now able to come in and add N-nucleotides to the existing P-nucleotides in a 5' to 3' direction that polymerases are known to function. On average 2-5 random base pairs are added to each 3' end generated after the action of the Artemis complex. The number of bases added is enough for the two newly synthesized ssDNA segments to undergo microhomology alignment during non-homologous end joining according to the normal Watson-Crick base pairing patterns (A-T, C-G). From there unpaired nucleotides are excised by an exonuclease, like the Artemis Complex (which has exonuclease activity in addition to endonuclease activity), and then template-dependent polymerases are able to fill the gaps, finally creating the new coding joint with the action of ligase to combine the segments. Although TdT does not discriminate among the four base pairs when adding them to the N-nucleotide segments, it has shown a bias for guanine and cytosine base pairs.[3]
Template Dependent Activity
edit
In a template-dependant manner, TdT can incorporate nucleotide across strand breaks in double-stranded DNA in a manner referred to as in trans in contrast to the in cis mechanism found in most polymerases. This occurs optimally with a one base-pair break between strands and less so with an increasing gap. This is facilitated by a subsection of TdT called Loop1 which selectively probes for short breaks in double-stranded DNA. Further, the discovery of this template dependant activity has lead to more convincing mechanistic hypotheses as to how the distribution of lengths of the additions of the N regions arise in V(D)J recombination[23].

Polymerase μ and polymerase λ exhibit similar in trans templated dependant synthetic activity to TdT, but without similar dependence on downstream double-stranded DNA[24]. Further, Polymerase λ has also been found to exhibit similar template-independent synthetic activity. Along with activity as a terminal transferase, it is known to also work in a more general template-dependent fashion.[25] The similatrities between TdT and polymerase μ suggest they are closely evolutionarily related[23].
Uses
editTerminal transferase has applications in molecular biology. It can be used in RACE to add nucleotides that can then be used as a template for a primer in subsequent PCR. It can also be used to add nucleotides labeled with radioactive isotopes, for example in the TUNEL assay (Terminal deoxynucleotidyl transferase dUTP Nick End Labeling) for the demonstration of apoptosis (which is marked, in part, by fragmented DNA). It is also used in the immunofluorescence assay for the diagnosis of acute lymphoblastic leukemia.[26]
In immunohistochemistry, antibodies to TdT can be used to demonstrate the presence of immature T and B cells and pluripotent hematopoietic stem cells, which possess the antigen, while mature lymphoid cells are always TdT-negative. While TdT-positive cells are found in small numbers in healthy lymph nodes and tonsils, the malignant cells of acute lymphoblastic leukaemia are also TdT-positive, and the antibody can, therefore, be used as part of a panel to diagnose this disease and to distinguish it from, for example, small cell tumours of childhood.[27]
TdT has also seen recent application in the De Novo synthesis of oligonucleotides, with TdT-dNTP tethered analogues capable of primer extension by 1 nt at a time.[28] In other words, the enzyme TdT has demonstrated the capability of making synthetic DNA by adding one letter at a time to a primer sequence.
See also
editReferences
edit- ^ Isobe M, Huebner K, Erikson J, Peterson RC, Bollum FJ, Chang LM, Croce CM (Sep 1985). "Chromosome localization of the gene for human terminal deoxynucleotidyltransferase to region 10q23-q25". Proceedings of the National Academy of Sciences of the United States of America. 82 (17): 5836–40. Bibcode:1985PNAS...82.5836I. doi:10.1073/pnas.82.17.5836. PMC 390648. PMID 3862101.
- ^ Yang-Feng TL, Landau NR, Baltimore D, Francke U (1986). "The terminal deoxynucleotidyltransferase gene is located on human chromosome 10 (10q23----q24) and on mouse chromosome 19". Cytogenetics and Cell Genetics. 43 (3–4): 121–6. doi:10.1159/000132309. PMID 3467897.
- ^ a b c Motea EA, Berdis AJ (May 2010). "Terminal deoxynucleotidyl transferase: the story of a misguided DNA polymerase". Biochimica et Biophysica Acta (BBA) - Proteins and Proteomics. 1804 (5): 1151–66. doi:10.1016/j.bbapap.2009.06.030. PMC 2846215. PMID 19596089.
- ^ Haeryfar SM, Hickman HD, Irvine KR, Tscharke DC, Bennink JR, Yewdell JW (Jul 2008). "Terminal deoxynucleotidyl transferase establishes and broadens antiviral CD8+ T cell immunodominance hierarchies". Journal of Immunology. 181 (1): 649–59. doi:10.4049/jimmunol.181.1.649. PMC 2587314. PMID 18566432.
- ^ BOLLUM FJ (1960). "Calf thymus polymerase". J Biol Chem. 235 (8): 2399–403. doi:10.1016/S0021-9258(18)64634-4. PMID 13802334.
- ^ Motea EA, Berdis AJ (2010). "Terminal deoxynucleotidyl transferase: the story of a misguided DNA polymerase". Biochim Biophys Acta. 1804 (5): 1151–66. doi:10.1016/j.bbapap.2009.06.030. PMC 2846215. PMID 19596089.
- ^ Gouge J, Rosario S, Romain F, Poitevin F, Béguin P, Delarue M (2015). "Structural basis for a novel mechanism of DNA bridging and alignment in eukaryotic DSB DNA repair". EMBO J. 34 (8): 1126–42. doi:10.15252/embj.201489643. PMC 4406656. PMID 25762590.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Loc'h J, Delarue M (2018). "Terminal deoxynucleotidyltransferase: the story of an untemplated DNA polymerase capable of DNA bridging and templated synthesis across strands". Curr Opin Struct Biol. 53: 22–31. doi:10.1016/j.sbi.2018.03.019. PMID 29656238. S2CID 4882661.
- ^ Hardy R (2008). "Chapter 7: B Lymphocyte Development and Biology". In Paul W (ed.). Fundamental Immunology (Book) (6th ed.). Philadelphia: Lippincott Williams & Wilkins. pp. 237–269. ISBN 978-0-7817-6519-0.
- ^ a b Ramadan K, Shevelev IV, Maga G, Hübscher U (2004). "De novo DNA synthesis by human DNA polymerase lambda, DNA polymerase mu and terminal deoxyribonucleotidyl transferase". J Mol Biol. 339 (2): 395–404. doi:10.1016/j.jmb.2004.03.056. PMID 15136041.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Bollum FJ, Chang LM, Tsiapalis CM, Dorson JW (1974). "Nucleotide polymerizing enzymes from calf thymus gland". Nucleic Acids and Protein Synthesis Part E. Methods in Enzymology. Vol. 29. pp. 70–81. doi:10.1016/0076-6879(74)29010-4. ISBN 9780121818920. PMID 4853390.
{{cite book}}: CS1 maint: multiple names: authors list (link) - ^ a b Chang LM, Bollum FJ (1970). "Doxynucleotide-polymerizing enzymes of calf thymus gland. IV. Inhibition of terminal deoxynucleotidyl transferase by metal ligands". Proc Natl Acad Sci U S A. 65 (4): 1041–8. Bibcode:1970PNAS...65.1041C. doi:10.1073/pnas.65.4.1041. PMC 283020. PMID 4985880.
- ^ a b c Deibel MR, Coleman MS (1980). "Biochemical properties of purified human terminal deoxynucleotidyltransferase". J Biol Chem. 255 (9): 4206–12. doi:10.1016/S0021-9258(19)85653-3. PMID 7372675.
- ^ Cherrier M, D'Andon MF, Rougeon F, Doyen N (Feb 2008). "Identification of a new cis-regulatory element of the terminal deoxynucleotidyl transferase gene in the 5' region of the murine locus". Molecular Immunology. 45 (4): 1009–17. doi:10.1016/j.molimm.2007.07.027. PMID 17854898.
- ^ Kubota T, Maezawa S, Koiwai K, Hayano T, Koiwai O (Aug 2007). "Identification of functional domains in TdIF1 and its inhibitory mechanism for TdT activity". Genes to Cells. 12 (8): 941–59. doi:10.1111/j.1365-2443.2007.01105.x. PMID 17663723. S2CID 25530793.
- ^ Zhang Y, Shi M, Wen Q, Luo W, Yang Z, Zhou M, Ma L (2012-01-01). "Antigenic stimulation induces recombination activating gene 1 and terminal deoxynucleotidyl transferase expression in a murine T-cell hybridoma". Cellular Immunology. 274 (1–2): 19–25. doi:10.1016/j.cellimm.2012.02.008. PMID 22464913.
- ^ Vashishtha AK, Wang J, Konigsberg WH (2016). "Different Divalent Cations Alter the Kinetics and Fidelity of DNA Polymerases". J Biol Chem. 291 (40): 20869–20875. doi:10.1074/jbc.R116.742494. PMC 5076500. PMID 27462081.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Delarue M, Boulé JB, Lescar J, Expert-Bezançon N, Jourdan N, Sukumar N; et al. (2002). "Crystal structures of a template-independent DNA polymerase: murine terminal deoxynucleotidyltransferase". EMBO J. 21 (3): 427–39. doi:10.1093/emboj/21.3.427. PMC 125842. PMID 11823435.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Steenberg ML, Lokhandwala MF, Jandhyala BS (1988). "Abnormalities in the sodium transport as the causative factor for enhanced norepinephrine overflow in the spontaneously hypertensive rat". Clin Exp Hypertens A. 10 (5): 833–41. doi:10.1080/07300077.1988.11878788. PMID 2846215.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ a b Schmoldt A, Benthe HF, Haberland G (1975). "Digitoxin metabolism by rat liver microsomes". Biochem Pharmacol. 24 (17): 1639–41. doi:10.1038/ni788. PMID doi.org/10.1038/ni788. S2CID 19412136.
{{cite journal}}: Check|pmid=value (help)CS1 maint: multiple names: authors list (link) - ^ Thai TH, Kearney JF (2004). "Distinct and opposite activities of human terminal deoxynucleotidyltransferase splice variants". J Immunol. 173 (6): 4009–19. doi:10.4049/jimmunol.173.6.4009. PMID 15356150. S2CID 40193319.
- ^ a b Thai TH, Kearney JF (2005). Isoforms of Terminal Deoxynucleotidyltransferase: Developmental Aspects and Function. Advances in Immunology. Vol. 86. pp. 113–36. doi:10.1016/S0065-2776(04)86003-6. ISBN 9780120044863. PMID 15705420.
- ^ a b Bland RD, Clarke TL, Harden LB (1976). "Rapid infusion of sodium bicarbonate and albumin into high-risk premature infants soon after birth: a controlled, prospective trial". Am J Obstet Gynecol. 124 (3): 263–7. doi:10.1016/0002-9378(76)90154-x. PMID j.jmb.2013.07.009.
{{cite journal}}: Check|pmid=value (help)CS1 maint: multiple names: authors list (link) - ^ Martin MJ, Blanco L (2014). "Decision-making during NHEJ: a network of interactions in human Pol μ implicated in substrate recognition and end-bridging". Nucleic Acids Res. 42 (12): 7923–34. doi:10.1093/nar/gku475. PMC 4081086. PMID 24878922.
- ^ Maga G, Ramadan K, Locatelli GA, Shevelev I, Spadari S, Hübscher U (Jan 2005). "DNA elongation by the human DNA polymerase lambda polymerase and terminal transferase activities are differentially coordinated by proliferating cell nuclear antigen and replication protein A". The Journal of Biological Chemistry. 280 (3): 1971–81. doi:10.1074/jbc.M411650200. PMID 15537631.
- ^ Faber J, Kantarjian H, Roberts MW, Keating M, Freireich E, Albitar M (Jan 2000). "Terminal deoxynucleotidyl transferase-negative acute lymphoblastic leukemia". Archives of Pathology & Laboratory Medicine. 124 (1): 92–7. doi:10.5858/2000-124-0092-TDTNAL. PMID 10629138.
- ^ Leong, Anthony S-Y; Cooper, Kumarason; Leong, F Joel W-M (2003). Manual of Diagnostic Cytology (2 ed.). Greenwich Medical Media, Ltd. pp. 413–414. ISBN 1-84110-100-1.
- ^ Palluk, S. et al. Nature Biotechnology 2018, 36, 645-650.
Further reading
edit- O'Malley DP, Orazi A (Aug 2006). "Terminal deoxynucleotidyl transferase-positive cells in spleen, appendix and branchial cleft cysts in pediatric patients". Haematologica. 91 (8): 1139–40. PMID 16885057.
- Yamashita N, Shimazaki N, Ibe S, Kaneko R, Tanabe A, Toyomoto T, Fujita K, Hasegawa T, Toji S, Tamai K, Yamamoto H, Koiwai O (Jul 2001). "Terminal deoxynucleotidyltransferase directly interacts with a novel nuclear protein that is homologous to p65". Genes to Cells. 6 (7): 641–52. doi:10.1046/j.1365-2443.2001.00449.x. PMID 11473582. S2CID 19573920.
- Chang LM, Bollum FJ (1986). "Molecular biology of terminal transferase". CRC Critical Reviews in Biochemistry. 21 (1): 27–52. doi:10.3109/10409238609113608. PMID 3524991.
- Maezawa S, Hayano T, Koiwai K, Fukushima R, Kouda K, Kubota T, Koiwai O (May 2008). "Bood POZ containing gene type 2 is a human counterpart of yeast Btb3p and promotes the degradation of terminal deoxynucleotidyltransferase". Genes to Cells. 13 (5): 439–57. doi:10.1111/j.1365-2443.2008.01179.x. PMID 18429817. S2CID 9698107.
- Taplin ME, Frantz ME, Canning C, Ritz J, Blumberg RS, Balk SP (Mar 1996). "Evidence against T-cell development in the adult human intestinal mucosa based upon lack of terminal deoxynucleotidyltransferase expression". Immunology. 87 (3): 402–7. doi:10.1046/j.1365-2567.1996.496571.x. PMC 1384108. PMID 8778025.
- Grupe A, Li Y, Rowland C, Nowotny P, Hinrichs AL, Smemo S, Kauwe JS, Maxwell TJ, Cherny S, Doil L, Tacey K, van Luchene R, Myers A, Wavrant-De Vrièze F, Kaleem M, Hollingworth P, Jehu L, Foy C, Archer N, Hamilton G, Holmans P, Morris CM, Catanese J, Sninsky J, White TJ, Powell J, Hardy J, O'Donovan M, Lovestone S, Jones L, Morris JC, Thal L, Owen M, Williams J, Goate A (Jan 2006). "A scan of chromosome 10 identifies a novel locus showing strong association with late-onset Alzheimer disease". American Journal of Human Genetics. 78 (1): 78–88. doi:10.1086/498851. PMC 1380225. PMID 16385451.
- Dworzak MN, Fritsch G, Fröschl G, Printz D, Gadner H (Nov 1998). "Four-color flow cytometric investigation of terminal deoxynucleotidyl transferase-positive lymphoid precursors in pediatric bone marrow: CD79a expression precedes CD19 in early B-cell ontogeny". Blood. 92 (9): 3203–9. doi:10.1182/blood.V92.9.3203. PMID 9787156.
- Fujita K, Shimazaki N, Ohta Y, Kubota T, Ibe S, Toji S, Tamai K, Fujisaki S, Hayano T, Koiwai O (Jun 2003). "Terminal deoxynucleotidyltransferase forms a ternary complex with a novel chromatin remodeling protein with 82 kDa and core histone". Genes to Cells. 8 (6): 559–71. doi:10.1046/j.1365-2443.2003.00656.x. PMID 12786946. S2CID 25223336.
- Kubota T, Maezawa S, Koiwai K, Hayano T, Koiwai O (Aug 2007). "Identification of functional domains in TdIF1 and its inhibitory mechanism for TdT activity". Genes to Cells. 12 (8): 941–59. doi:10.1111/j.1365-2443.2007.01105.x. PMID 17663723. S2CID 25530793.
- Suzuki Y, Yoshitomo-Nakagawa K, Maruyama K, Suyama A, Sugano S (Oct 1997). "Construction and characterization of a full length-enriched and a 5'-end-enriched cDNA library". Gene. 200 (1–2): 149–56. doi:10.1016/S0378-1119(97)00411-3. PMID 9373149.
- Bridges SL (Aug 1998). "Frequent N addition and clonal relatedness among immunoglobulin lambda light chains expressed in rheumatoid arthritis synovia and PBL, and the influence of V lambda gene segment utilization on CDR3 length". Molecular Medicine. 4 (8): 525–53. doi:10.1007/BF03401757. PMC 2230400. PMID 9742508.
- Liu L, McGavran L, Lovell MA, Wei Q, Jamieson BA, Williams SA, Dirks NN, Danielson MS, Dubie LM, Liang X (Jun 2004). "Nonpositive terminal deoxynucleotidyl transferase in pediatric precursor B-lymphoblastic leukemia". American Journal of Clinical Pathology. 121 (6): 810–5. doi:10.1309/QD18-PPV1-NH3T-EUTF. PMID 15198352.
- Yang B, Gathy KN, Coleman MS (Apr 1994). "Mutational analysis of residues in the nucleotide binding domain of human terminal deoxynucleotidyl transferase". The Journal of Biological Chemistry. 269 (16): 11859–68. doi:10.1016/S0021-9258(17)32652-2. PMID 8163485.
- Thai TH, Kearney JF (Sep 2004). "Distinct and opposite activities of human terminal deoxynucleotidyltransferase splice variants". Journal of Immunology. 173 (6): 4009–19. doi:10.4049/jimmunol.173.6.4009. PMID 15356150. S2CID 40193319.
- Shimazaki N, Fujita K, Koiwai O (Mar 2002). "[Expression and function of terminal deoxynucleotidyl-transferase and discovery of novel DNA polymerase mu]". Seikagaku. The Journal of Japanese Biochemical Society. 74 (3): 227–32. PMID 11974916.
- Mahajan KN, Mitchell BS (Sep 2003). "Role of human Pso4 in mammalian DNA repair and association with terminal deoxynucleotidyl transferase". Proceedings of the National Academy of Sciences of the United States of America. 100 (19): 10746–51. Bibcode:2003PNAS..10010746M. doi:10.1073/pnas.1631060100. PMC 196874. PMID 12960389.
- Mahajan KN, Gangi-Peterson L, Sorscher DH, Wang J, Gathy KN, Mahajan NP, Reeves WH, Mitchell BS (Nov 1999). "Association of terminal deoxynucleotidyl transferase with Ku". Proceedings of the National Academy of Sciences of the United States of America. 96 (24): 13926–31. Bibcode:1999PNAS...9613926M. doi:10.1073/pnas.96.24.13926. PMC 24167. PMID 10570175.
- Ibe S, Fujita K, Toyomoto T, Shimazaki N, Kaneko R, Tanabe A, Takebe I, Kuroda S, Kobayashi T, Toji S, Tamai K, Yamamoto H, Koiwai O (Sep 2001). "Terminal deoxynucleotidyltransferase is negatively regulated by direct interaction with proliferating cell nuclear antigen". Genes to Cells. 6 (9): 815–24. doi:10.1046/j.1365-2443.2001.00460.x. PMID 11554927. S2CID 19287230.
External links
edit- Terminal+Deoxyribonucleotidyltransferase at the U.S. National Library of Medicine Medical Subject Headings (MeSH)